
INLEIDING
Chronische pijn is een complexe en vaak onbegrepen aandoening, die zich op zeer uiteenlopende manieren manifesteert en sterk verschilt van persoon tot persoon. Er is sprake van chronische pijn wanneer pijn langer dan drie maanden aanhoudt, langer dan de normale genezingsduur.
Ze kan verband houden met een vastgestelde aandoening– zoals een neuropathie, een ontsteking of kanker – maar soms blijft ze zonder duidelijke organische oorzaak.
Vanuit haar multidimensionele oorsprong is chronische pijn het resultaat van complexe pathofysiologische mechanismen, vaak verweven met psychologische en sociale factoren. Deze complexiteit bemoeilijkt niet alleen de diagnose, maar ook de behandeling, die een globale, geïndividualiseerde en vaak multidisciplinaire aanpak vereist, gericht op de werkelijkheid die elke patiënt ervaart.
Vaak doorlopen de betrokkenen een therapeutische dwaaltocht, temeer omdat bepaalde soorten chronische pijn andere aandoeningen nabootsen of daarmee verward kunnen worden, waardoor de herkenning en behandeling ervan moeilijker worden. Deze realiteit maakt het vandaag de dag tot een echte uitdaging voor de volksgezondheid.
Om dit fenomeen, dat nog te weinig in al zijn rijkdom en complexiteit wordt begrepen, te verduidelijken, zullen we samen de laatste wetenschappelijke ontwikkelingen op dit gebied bekijken, om een beter inzicht te krijgen in de soorten chronische pijn, hun multifactoriële oorzaken, en de natuurlijke, integratieve protocollen die een doeltreffende aanpak mogelijk maken.
Soorten pijn


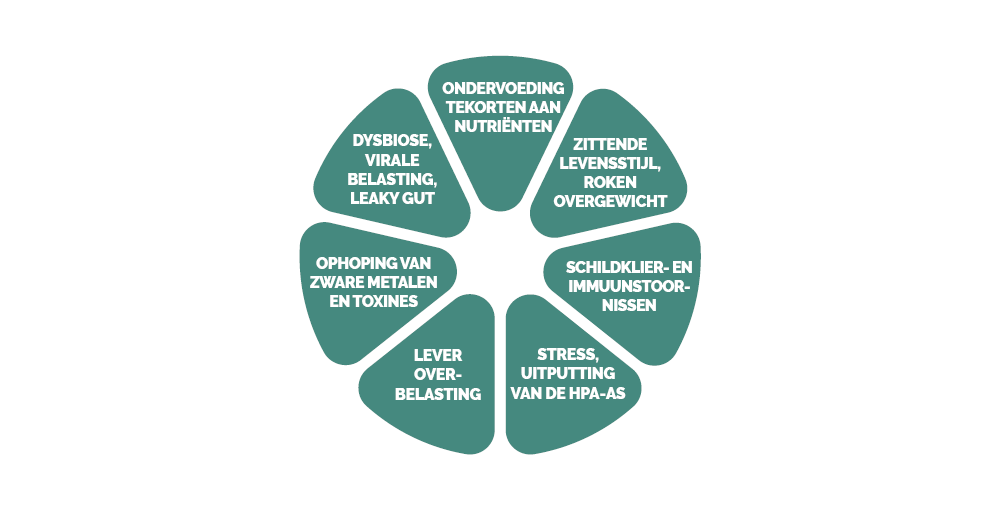
De verschillende multifactoriële oorzaken
Chronische pijn is een complexe problematiek, waarvan de oorzaken en verergerende factoren vaak meervoudig en met elkaar verweven zijn. Ze laat zich niet herleiden tot een eenvoudig letsel of een op zichzelf staand onevenwicht, maar is eerder het resultaat van een geheel van biologische, psychologische en omgevingsfactoren die met elkaar in wisselwerking staan.
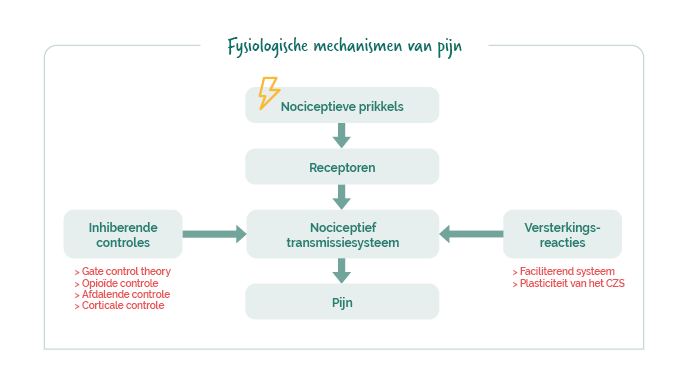
De fysiologische mechanismen van pijn
Pijn begint met een nociceptieve prikkel (weefselbeschadiging, ontsteking, uitrekking, enz.) die wordt opgevangen door perifere nociceptoren. Dit zijn vrije zenuwuiteinden die specifieke ionkanalen tot expressie brengen, gevoelig voor mechanische, thermische of chemische prikkels.
De activatie van deze receptoren wekt een actiepotentiaal op dat via afferente vezels (A-delta en C) wordt doorgegeven tot in de dorsale hoorn van het ruggenmerg. Het signaal wordt vervolgens verwerkt en doorgegeven via het nociceptieve transmissiesysteem, dat de zenuwbanen van de periferie tot aan de hersenen omvat. Op dat moment wordt de pijn daadwerkelijk waargenomen.
CHRONISCHE PIJN EN ONTSTEKING
Bevorderende factoren
Laaggradige ontsteking en neuro-inflammatie
Chronische laaggradige ontsteking wordt gekenmerkt door een langdurige maar matige activering van het aangeboren immuunsysteem, zonder de klassieke klinische symptomen van acute ontsteking (warmte, roodheid, oedeem). Het weerspiegelt een verhoogde alarmtoestand van het lichaam die vaak onopgemerkt blijft of niet wordt gehoord. Deze toestand speelt een centrale rol bij tal van ziekten die chronische pijn veroorzaken. Immuuncellen geven ontstekingsmediatoren af die de perifere en centrale neuronen sensibiliseren, waardoor de pijnoverdracht wordt verstoord. Ze kunnen ook het ruggenmerg en de dorsale ganglia binnendringen. Deze langdurige ontstekingsactivatie beïnvloedt bovendien stemming en gedrag en wordt in verband gebracht met verschillende psychiatrische stoornissen, waaronder stemmingsstoornissen, verslavingen, schizofrenie, ADHD en autisme (1).
Daarbij is het belangrijk op te merken dat roken, alcoholgebruik, obesitas, een medische voorgeschiedenis van chronische pijn en psychologische factoren (zoals stress en angst) allemaal belangrijke elementen zijn die dit ontstekingsproces in stand houden.
Darmdysbiose, leverfunctie en chronische pijn
Darmdysbiose, gekenmerkt door een verstoring van het evenwicht in het microbioom, bevordert een toegenomen darmpermeabiliteit (« leaky gut ») waardoor lipopolysacchariden (LPS) en andere microbiële toxines in de systemische bloedsomloop terechtkomen (2). Deze vetoplosbare verbindingen moeten door delever worden omgezet via de fase I- en fase II-ontgifting om te worden omgezet in wateroplosbare moleculen die kunnen worden uitgescheiden. Wanneer deze capaciteit overschreden wordt, ontstaat er een systemische toxische opstapeling, die vooral weefsels met een hoog lipidengehalte aantast, zoals de hersenen (ongeveer 60% lipiden), wat bijdraagt aan neurotoxiciteit (3).
Centrale rol van de lever bij pijnmodulatie
Darmdysbiose en leveroverbelasting leiden tot een aanhoudende activering van het immuunsysteem, waardoor pro-inflammatoire cytokines (TNF-α, IL-6, IL-1β) vrijkomen die de bloed-hersenbarrière passeren en de microgliale cellen in de hersenen activeren (4). Dit fenomeen staat centraal in de mechanismen van chronische neuro-inflammatie die verantwoordelijk zijn voor centrale pijnsensibilisatie, een belangrijk proces bij neuropathische en chronische pijn (5).
De lever speelt een sleutelrol in het voorkomen van neurotoxemie door deze in vet oplosbare toxines te neutraliseren. Het ondersteunen van de leverdetoxificatie, met name fase II (conjugatie), is essentieel om de systemische toxische belasting en perifere ontsteking te verminderen die de neuropathische pijn onderhouden (6).
Een tekort aan glutathion, de belangrijkste cellulaire antioxidant en een belangrijke cofactor in fase II, versterken de neurotoxische impact door de capaciteit tot neutralisatie van vrije radicalen en toxische metabolieten te verminderen (7).
- In het schema zien we de verschillende fasen van de leverdetoxificatie en de micronutriënten die de activering van deze fasen ondersteunen.
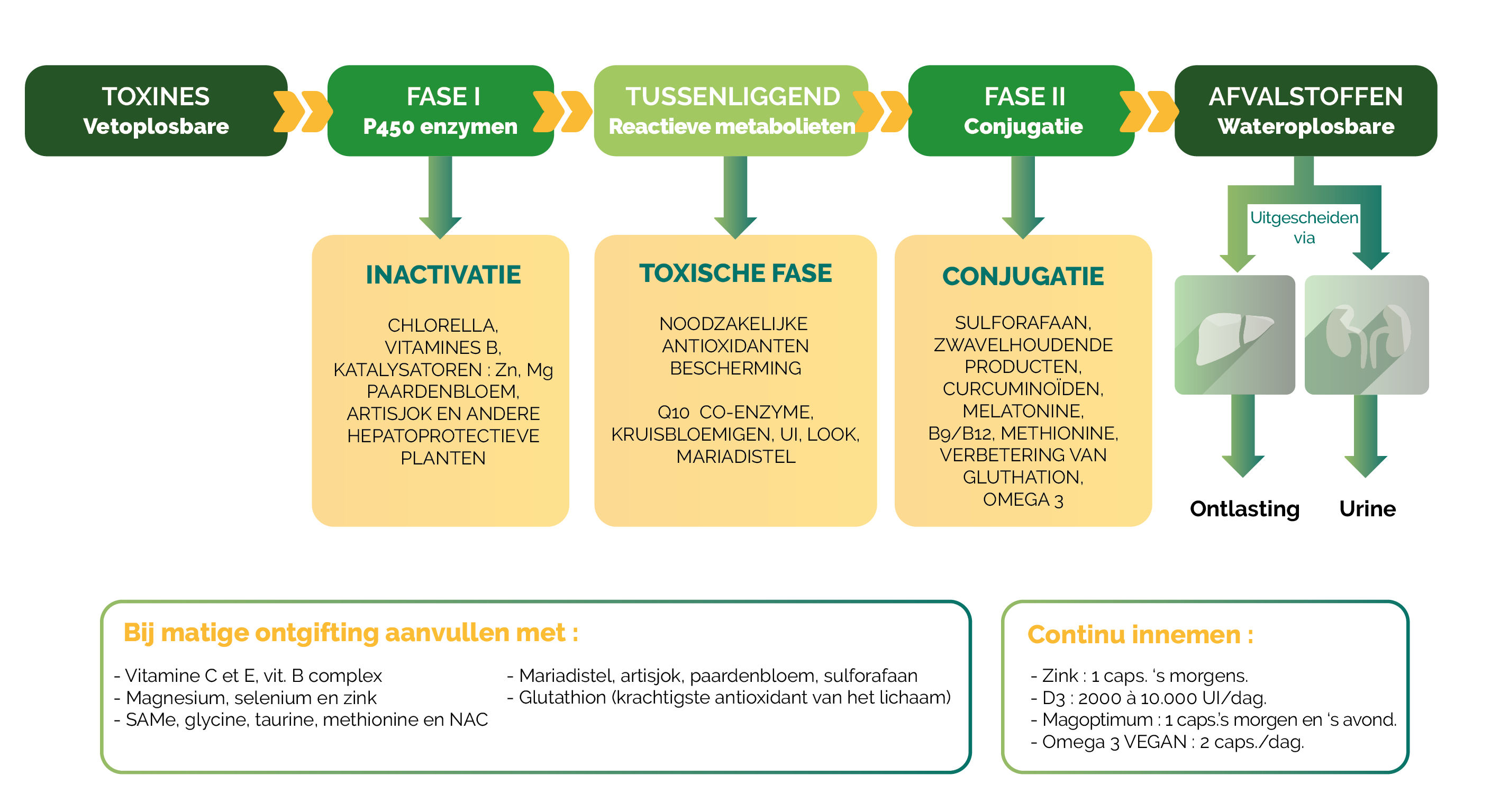
Door de leverdetoxificatie te versterken, dragen deze micronutriënten bij tot het verminderen van neuro-inflammatie en centrale sensitisatie, en bieden ze een relevante therapeutische invalshoek bij de aanpak van chronische neuropathische pijn.
Stress en uitputting van de HPA-as
Langdurige stress leidt tot een aanhoudende activatie van de hypothalamus-hypofyse-bijnier-as (HPA), met een overproductie van cortisol, gevolgd door een geleidelijke uitputting van het systeem. Dit resulteert in een gedysreguleerde HPA-respons (8). Deze ontregeling draagt bij tot de sensitisatie van pijncircuits en bevordert de overgang naar chronische pijn.
Op lange termijn verstoren hoge cortisolgehalten (9) de synaptische plasticiteit, vooral in de prefrontale cortex en de hippocampus. Deze veranderingen veroorzaken een hyperprikkelbaarheid van de dorsale hoorn van het ruggenmerg en een centralisatie van pijn, waardoor de pijnperceptiesystemen gevoeliger worden, zelfs voor lichte stimuli. Tegelijkertijd stimuleert stress de afgifte van pro-inflammatoire cytokinen, zoals IL-6 en TNF-α, die de bloed-hersenbarrière kunnen passeren (10). Deze cytokinen activeren de microgliacellen in het centrale zenuwstelsel, waardoor een toestand van aanhoudende neuro-inflammatie in stand wordt gehouden (11).
Deze langdurige microglia-activatie beïnvloedt de overdracht van nociceptieve signalen, en draagt zo bij aan het behoud van een verhoogde pijnwaakzaamheid.
Bovendien verstoort chronische stress het tryptofaanmetabolisme door de activiteit van het enzym IDO (indoleamine-2,3-dioxygenase)te verhogen, waardoor tryptofaan wordt afgeleid van de serotonerge route naar de productie van neuroactieve kynurenines. Deze metabolieten kunnen de NMDA-receptoren (N-Méthyl-D-Aspartaat) activeren, wat de neuronale excitatie en excitotoxiciteit versterkt, en zo de waargenomen pijn verergert (12)(13).
Klinische gevolgen zijn onder meer : slaapstoornissen, chronische vermoeidheid, hoofdpijn, stressovergevoeligheid en cognitieve achteruitgang, die vaak voorkomen bij aandoeningen zoals fibromyalgie en neuropathie (14).
Zo ontstaat een vizieuse cirkel waarin stress en pijn elkaar wederzijds versterken: stress bevordert ontsteking en pijn, terwijl chronische pijn de perceptie van stress verhoogt, via dezelfde neuro-endocriene en immuunroutes. Deze tweezijdige interactie tussen pijn en ontsteking maakt het zonder een globale en multidisciplinaire aanpak des te moeilijker om uit dit proces te komen.

Een gevestigde tweezijdige interactie
Laaggradige ontsteking werkt als een voedingsbodem voor chronische pijn, door een sensitisatie van de centrale en perifere nociceptieve routes in stand te houden.
Klinisch en experimenteel onderzoek heeft aangetoond dat:
- Klinisch en experimenteel onderzoek heeft aangetoond dat: – Bij patiënten met chronische musculoskeletale pijn werden verhoogde concentraties van pro-inflammatoire cytokines, in het bijzonder IL-6 en TNF-α, in het bloed gecorreleerd met de waargenomen pijnintensiteit (15).
- Bij fibromyalgie — beschouwd als een prototype van nociplastische pijn — bleek uit een meta-analyse dat bepaalde ontstekingsmediatoren significant verhoogd waren, wat suggereert dat zelfs bij afwezigheid van ontsteking de pijn kan worden verergerd door de aanwezigheid van ontstekingsmediatoren. (16).
- Neuro-imagingstudies hebben de activatie van microglia in de hersenen aangetoond in een context van chronische perifere ontsteking. Deze gliale activatie draagt bij aan neuro-inflammatie en centrale sensitisatie, sleutelmechanismen in de overgang naar chronische pijn (17).
- Op neuro-immunologisch vlak, zijn cytokinen zoals IL-1β en TNF-α betrokken bij de verstoring van de zenuwgeleiding en de verhoogde prikkelbaarheid van nociceptieve vezels, met name na een zenuwbeschadiging (18).
- Ten slotte, kan een ontregeling van het tryptofaanmetabolisme, onder invloed van een overmatige activatie van NF-κB en in een ontstekingscontext, leiden tot een afleiding naar de kynurenine-route ten koste van de serotoninesynthese. Bepaalde metabolieten die uit deze route voortkomen, zoals kinolinezuur, activeren de NMDA-receptor (N-Méthyl-D-Aspartaat), waardoor hyperalgesie en sensitisatie worden versterkt (19). Deze metabolische omleiding maakt het gebruik van tryptofaan bij bepaalde nociplastische pijnen delicaat, omdat het hyperalgetisch kan worden, vooral als de ontsteking niet onder controle is.
De belangrijkste indicatoren voor een ontstekingsproces
Tot op heden bestaat er geen unieke biomarker die formeel een chronische laaggradige ontsteking kan diagnosticeren. Verschillende biologische indicatoren, in combinatie met de anamnese en het klinische beeld, kunnen echter het vermoeden van een stille ontsteking versterken. Deze indicatoren wijzen op een aanhoudende activering van het aangeboren immuunsysteem en een verstoring van de metabolische en neuro-immuunregulatie.
1. Ultrasensitieve CRP (CRPus)
In tegenstelling tot standaard CRP maakt ultrasensitieve CRP het mogelijk om lage verhogingen (0,3 tot 10 mg/L), te detecteren, die typisch zijn voor stille ontstekingen (20).
- < 1 mg/L : laag risico (minieme ontsteking)
- 1–3 mg/L : matige ontsteking
- 3 mg/L : aanzienlijke laaggradige ontsteking
2. Elektroforese van plasma-eiwitten (EPP)
Maakt het mogelijk subtiele veranderingen in de eiwitfracties op te sporen (21).
- ↗︎ Alpha-1-globulines (bv: α1-antitrypsine, orosomucoïde): kenmerkend voor de acute ontstekingsfase.
- ↗︎ Alpha-2-globulines (haptoglobine, α2-macroglobuline) et ↘︎ Albumine: kenmerkend voor een chronische ontsteking
3. Vetzuurstatus (omega-6/omega-3 verhouding)
Een verstoorde verhouding tussen AA/EPA (AA: arachidonzuur) of omega-6/omega-3 (>10) wordt vaak in verband gebracht met een chronische ontstekingsreactie. Een verhouding > 10 wordt gekoppeld aan een pro-inflammatoire activering van het vetmetabolisme en een stimulatie van de arachidonzuur afgeleide routes (PGE2, LTB4…) (22).
4. Serumferritine
Wanneer ferritine matig verhoogd is bij een normale of lage ijzerverzadiging, kan dit wijzen op een chronische ontstekingsaandoening (behalve hemochromatose) (23).
Wijzigingen in verschillende biologische parameters tijdens een inflammatoire ijzertekort:
- Hemoglobine – In daling ↘︎
- Ijzer – In daling ↘︎
- Verzadigingsgraad – Normaal of laag ↘︎
- Ferritine – Normaal of hoog ↗︎
Bron: https://www.revmed.ch/revue-medicale-suisse/2011/revue-medicale-suisse-313/anemie-ferriprive-inflammatoire-ou-mixte-comment-orienter-le-diagnostic
5. Homocysteïne
Een verhoogd homocysteïnegehalte wordt in verband gebracht met cardiovasculaire risico’s, maar volgens sommige studies ook met activering van microglia en neuropathische pijn. Het is ook belangrijk om de dosering van actief B12 en erytrocytair B9 te controleren bij het doseren van homocysteïne, omdat een verhoogd homocysteïnegehalte verband kan houden met een tekort aan B12 en B9 (24).
6. Zonuline
Zonuline, een regulator van de darm-doorlaatbaarheid, is een onrechtstreekse marker voor de darmdoorlaatbaarheid. Een verhoging bevordert de doorgang van LPS en bacteriële fragmenten naar de systemische circulatie, waarbij TLR4 (Toll-Like Receptor 4) wordt gestimuleerd (25).
7. LPS (Lipopolysacchariden) et LBP (LPS-binding protein)
- PS is een bestanddeel van Gram− bacteriën. Wanneer het in de circulatie terechtkomt, stimuleert het TLR4 en bijgevolg de ontstekingscascade (26).
- is een levereiwit dat zich bindt aan LPS: de concentratie stijgt mee met de circulerende LPS-last (27).

- Ultrasensitieve CRP (CRPus)
- Plasma-eiwitelektroforese (EPP)
- Vetzurenstatus
- Serumferritine
- Homocysteïne
- Zonuline
- LPS en LBP
WELKE OPLOSSINGEN KUNNEN WORDEN OVERWOGEN
Het vermogen van PEA
PEA is een natuurlijk endogeen lipide dat onmiddellijk en lokaal door het lichaam wordt aangemaakt als reactie op bepaalde prikkels: ontstekingen, infecties, intoxicatie, verwondingen, trauma’s en stress. Wetenschappers hebben vastgesteld dat het in grote hoeveelheden wordt aangemaakt in weefsels die degeneratie of letsels vertonen, dus in geval van ontstekingen.
PEA is een vorm van weefselbescherming die helpt bij weefselherstel. Het heeft een endocannabinoïde-achtig effect, maar is niet psychotroop, waardoor het de pijnperceptie reguleert. PEA zou ook een rol kunnen spelen bij de desensibilisatie van warmtereceptoren, die bij ontstekingen overactief zijn.
Dit endogene, pijnstillende en ontstekingsremmende lipide is bijzonder nuttig bij neuropathische pijn door modulatie van microglia en activering van PPAR-α (Peroxisome Proliferator-Activated Receptor alpha).
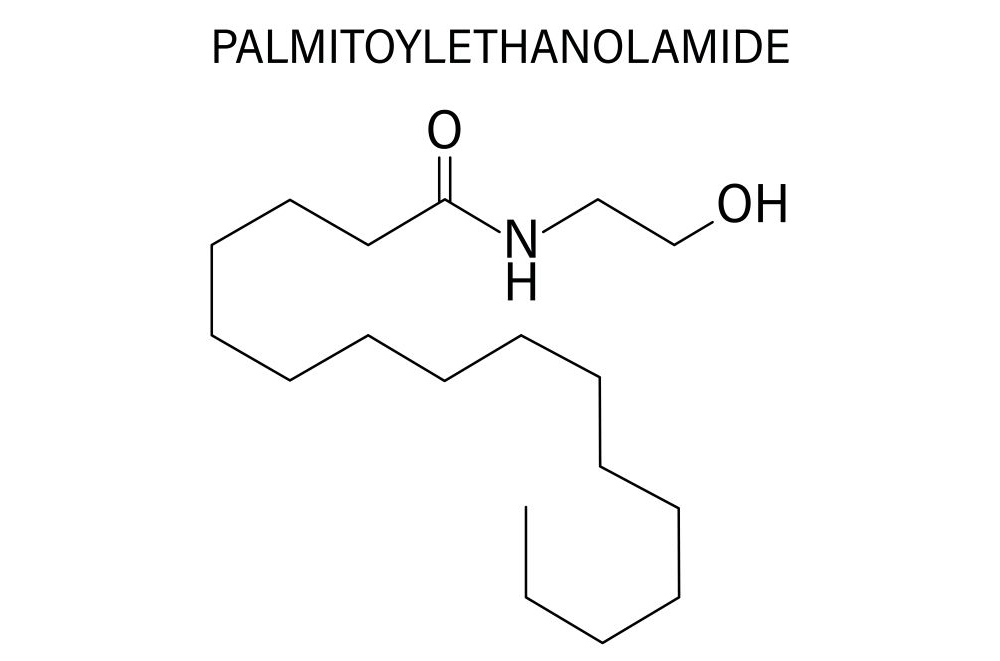
Een systematische review en meta-analyse, uitgevoerd in 2023, evalueerde de werkzaamheid van PEA bij de behandeling van chronische pijn. Van de 253 geïdentificeerde artikelen werden 11 gerandomiseerde dubbelblinde studies opgenomen, met in totaal 774 patiënten. De resultaten tonen aan dat PEA de pijnscores significant vermindert in vergelijking met placebo of andere behandelingen. Er werden ook verbeteringen in de levenskwaliteit en motorische functie gemeld, zonder ernstige bijwerkingen of medicamenteuze interacties (28).
Een andere gerandomiseerde, dubbelblinde, placebogecontroleerde studie evalueerde de werkingsmechanismen van PEA bij 14 gezonde vrijwilligers. Na 4 weken PEA-inname (3 × 400 mg/dag) werden verschillende pijnmarkers gemeten. De resultaten tonen een significante afname van de geïnduceerde pijn, een betere tolerantie voor koude en druk, evenals een gunstige modulatie van pijn (vermindering van allodynie). Deze gegevens wijzen erop dat PEA klinisch relevante pijnstillende effecten heeft via zowel perifere, centrale als modulerende mechanismen, met name in het kader van nociplastische pijn (29).
Het vermogen van Gember
Gember bevat meer dan 400 natuurlijke verbindingen, waarvan gingerolen, shogaolen en paradolen het meest zijn bestudeerd om hun ontstekingsremmende en pijnstillende effecten.
Vooral 6-gingerol is het belangrijkste actieve bestanddeel van verse gember, terwijl shogaolen, die tijdens het drogen worden gevormd, krachtiger zijn wat betreft hun therapeutische werking.
Deze moleculen hebben een modulerende werking op verschillende ontstekingsroutes, met name door de productie van prostaglandinen en pro-inflammatoire cytokines (TNF-α, IL-1β, IL-6), te remmen, belangrijke mediatoren bij chronische pijn en neuro-inflammatie.

Werkingsmechanismen die betrokken zijn bij pijnmodulatie:
Gember werkt als een niet-selectieve remmer van de cyclo-oxygenase (COX-1 et COX-2) en lipoxygenase (5-LOX) routes, waardoor de productie van prostaglandinen (PGE2) en leukotriënen die betrokken zijn bij ontstekingen en nociceptie wordt verminderd (30). Deze werking is vergelijkbaar met die van bepaalde NSAID’s, maar met een betere gastro-intestinale tolerantie.
Bovendien remmen gingerolen de nucleaire translocatie van NF‑κB (nucleaire kappa B van geactiveerde B-lymfocyten), een belangrijke factor bij de activering van pro-inflammatoire genen, wat leidt tot een afname van de productie van TNF-α, IL‑1β en IL‑6. Deze werking draagt bij aan het verminderen van neuro-inflammatie die betrokken is bij het chronisch worden van pijn (31).
De fenolverbindingen in gember kunnen de bloed-hersenbarrière passeren en een wisselwerking aangaan met de immuuncellen in de hersenen, met name de microglia. Preklinische studies suggereren dat gember een ontstekingsremmend fenotype (M2) van microgliale cellen bevordert, wat bijdraagt aan een neuroprotectief effect (32).
De pijnstillende werking wordt verklaard door een remming van het enzym COX-2 (cyclo-oxygenase-2), dat betrokken is bij ontstekingen, maar ook door een regulerend effect op de TRPV1-pijnreceptoren (Transient Receptor Potential Vanilloid 1) die bij chronische ontstekingen vaak overactief zijn. Het zijn dezelfde receptoren waarop capsaïcine zich richt, dat wordt gebruikt om de overdracht van pijnsignalen in de perifere zenuwen te verminderen (33).
Ten slotte vermindert gember oxidatieve stress door de activiteit van de antioxidante enzymen SOD (SuperOxyde Dismutase) en GPx (Glutathione Peroxydase), te verhogen, waardoor neuronen en perifere weefsels worden beschermd tegen schade door vrije radicalen, die vaak voorkomen bij chronische ontstekingen. (34).
Het vermogen van Krill
Krillolie onderscheidt zich door zijn unieke samenstelling aan omega-3-vetzuren (EPA en DHA) die gebonden zijn aan fosfolipiden, in tegenstelling tot visolie, waar ze voornamelijk in triglyceridevorm voorkomen. Deze fosfolipiden verhogen de biologische beschikbaarheid, in het bijzonder in neuronale en immuuncelmembranen, wat belangrijk is bij chronische pijn van inflammatoire of neuropathische oorsprong. Krill bevat ook astaxanthine, een krachtige antioxidant met neuroprotectieve eigenschappen.
De pijnstillende en ontstekingsremmende eigenschappen van krill zijn gebaseerd op verschillende complementaire mechanismen. Het vermindert de productie van pro-inflammatoire cytokines zoals TNF-α en l’IL-6, die betrokken zijn bij het ontstaan en voortduren van chronische pijn. Deze modulatie beperkt de sensibilisatie van nociceptoren en overmatige neuronale prikkelbaarheid.
Op centraal niveau hebben de omega-3-vetzuren in krill een remmende werking op de overmatige activering van microglia waardoor neuro-inflammatie en centrale sensibilisatie worden verminderd. Tegelijkertijd ondersteunt krill het energetisch metabolisme van neuronen door de mitochondriale functie te verbeteren, wat bijdraagt tot het verminderen van vermoeidheid en aanhoudende pijnprikkels. Astaxanthine werkt synergetisch, beperkt lipideperoxidatie en beschermt de neuronale membranen.

Recente studies bevestigen dat omega-3-vetzuren uit de zee, met name die uit krill, een gunstige invloed hebben op de ontstekings- en oxidatieve processen die gepaard gaan met aanhoudende pijn. Door tegelijkertijd in te werken op perifere en centrale mediatoren, profileert krill zich als een veelbelovende natuurlijke optie in een integratieve aanpak van chronische pijn (35).
Het vermogen van Fycocyanine
Fycocyanine is een natuurlijk blauw pigment-eiwit dat voorkomt in cyanobacteriën zoals spirulina. Het heeft krachtige antioxydanten, ontstekingsremmende en immunomodulerende eigenschappen. Zijn unieke bioactieve profiel, met in het bijzonder zijn tetramere structuur die rijk is aan chromoforen, stelt hem in staat om effectief in te werken op vrije radicalen en ontstekingsmediatoren.
Bij chronische pijn remt het de NF-κB-signaalroute, wat leidt tot een vermindering van de expressie van pro-ontstekingsbevorderende cytokines zoals l’IL-1β, l’IL-6 en TNF-α. Het moduleert ook de activiteit van COX-2 (Cyclo-Oxygenase-2), waardoor de productie van pijnveroorzakende prostaglandinen wordt beperkt, zonder de ongewenste gastro-intestinale effecten die gepaard gaan met conventionele ontstekingsremmers.
Op neuroprotectief vlak, vermindert fycocyanine oxidatieve stress door lipideperoxidatie te remmen, waardoor de integriteit van neuronale membranen behouden blijft, wat vooral relevant is bij neuropathische pijn. Bovendien wijzen preklinische modellen erop dat het de endogene antinociceptieve routes versterkt, met in het bijzonder via de activering van de GABAergische en opioïde systemen, waardoor de centrale pijnstillende werking wordt versterkt.

Experimentele studies hebben aangetoond dat fycocyanine pijnstillende effecten heeft die vergelijkbaar zijn met die van paracetamol of NSAID’s (niet-steroïde ontstekingsremmers), maar met een uitstekend tolerantieprofiel, zonder noemenswaardige effecten op de lever of de spijsvertering. (36, 37).
Het vermogen van Saffraan
Saffraan, rijk aan bioactieve verbindingen zoals crocine, safranal en picrocrocine, bezit goed gedocumenteerde pijnstillende, ontstekingsremmende en neuromodulerende eigenschappen. Deze moleculen werken op het kruispunt van neuro-ontstekings- en neurotransmissieroutes, waardoor het bijzonder geschikt is voor chronische pijn van gemengde oorsprong (nociceptief, neuropathisch, nociplastisch).
Op ontstekings vlak moduleert saffraan de NF-κB-signaalroute, wat leidt tot een remming van de productie van pro-inflammatoire cytokinen zoals IL-6 en TNF-α, die betrokken zijn bij perifere en centrale sensibilisatie. Het oefent tevens een antioxidatieve werking in de hersenen uit, waardoor neuronale oxidatieve stress – een belangrijke factor in het ontstaan van gecentraliseerde chronische pijn – wordt beperkt.
Bovendien werkt saffraan in op het serotoninerge,dopaminerge en opioïde systeem, wat bijdraagt aan een verhoging van de pijngrens en een verbetering van de nociceptieve tolerantie. De werking op de neurotransmissie hangt ook samen met positieve effecten op de stemming, die vaak verandert bij chronische pijnsyndromen. Deze modulatie gebeurt vooral door regulering van de tryptofaanroute en remming van monoamineoxidase (MAO), waardoor het antidepressieve effect wordt versterkt.
Klinische studies hebben de doeltreffendheid van saffraan bevestigd bij het verminderen van pijn bij verschillende chronische aandoeningen, zoals dysmenorroe, fibromyalgie of lage rugpijn, en als aanvullende ondersteuning bij depressieve toestanden (38, 39).

NATUURLIJKE AAPAK
Suggesties voor protocollen
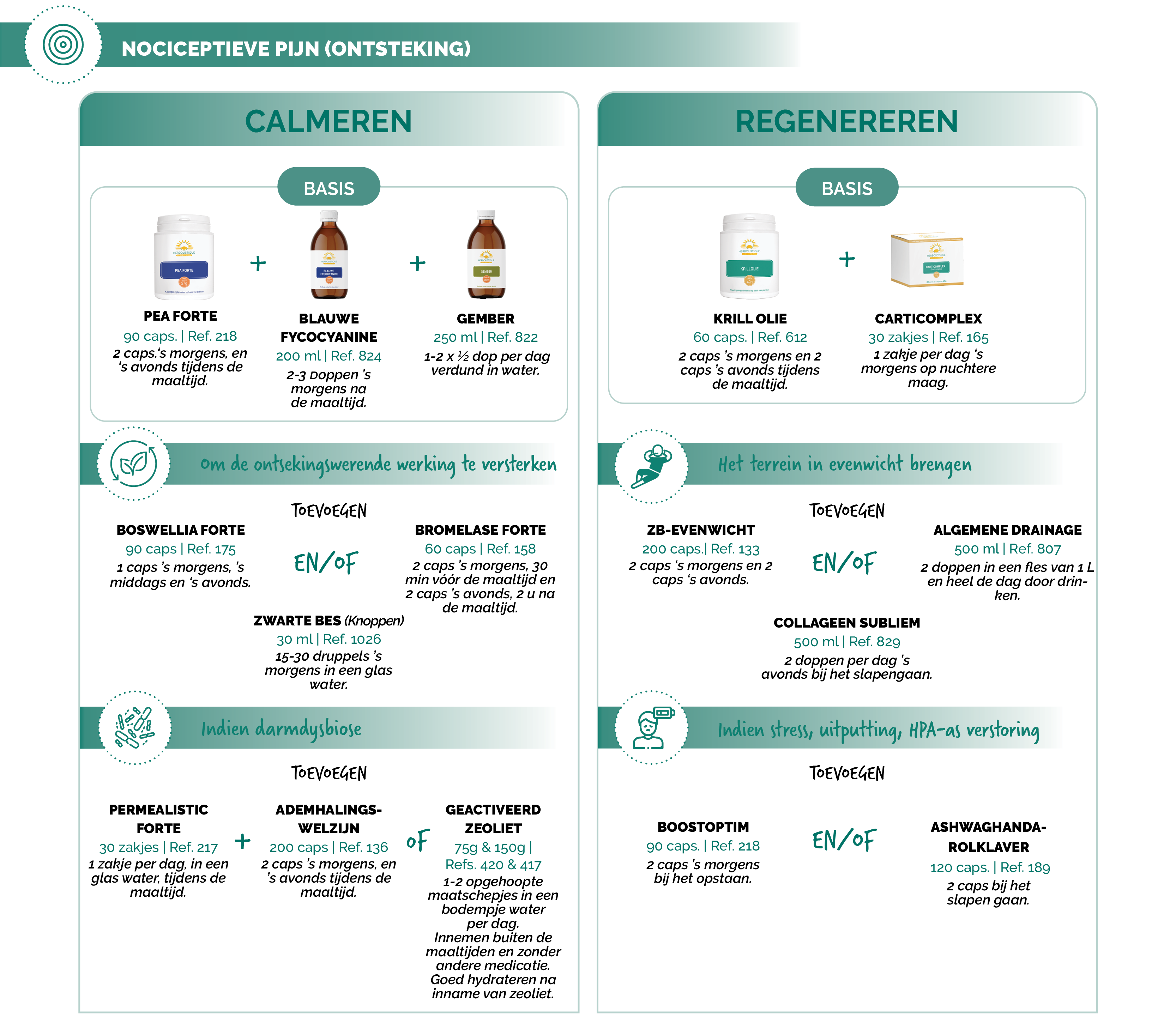
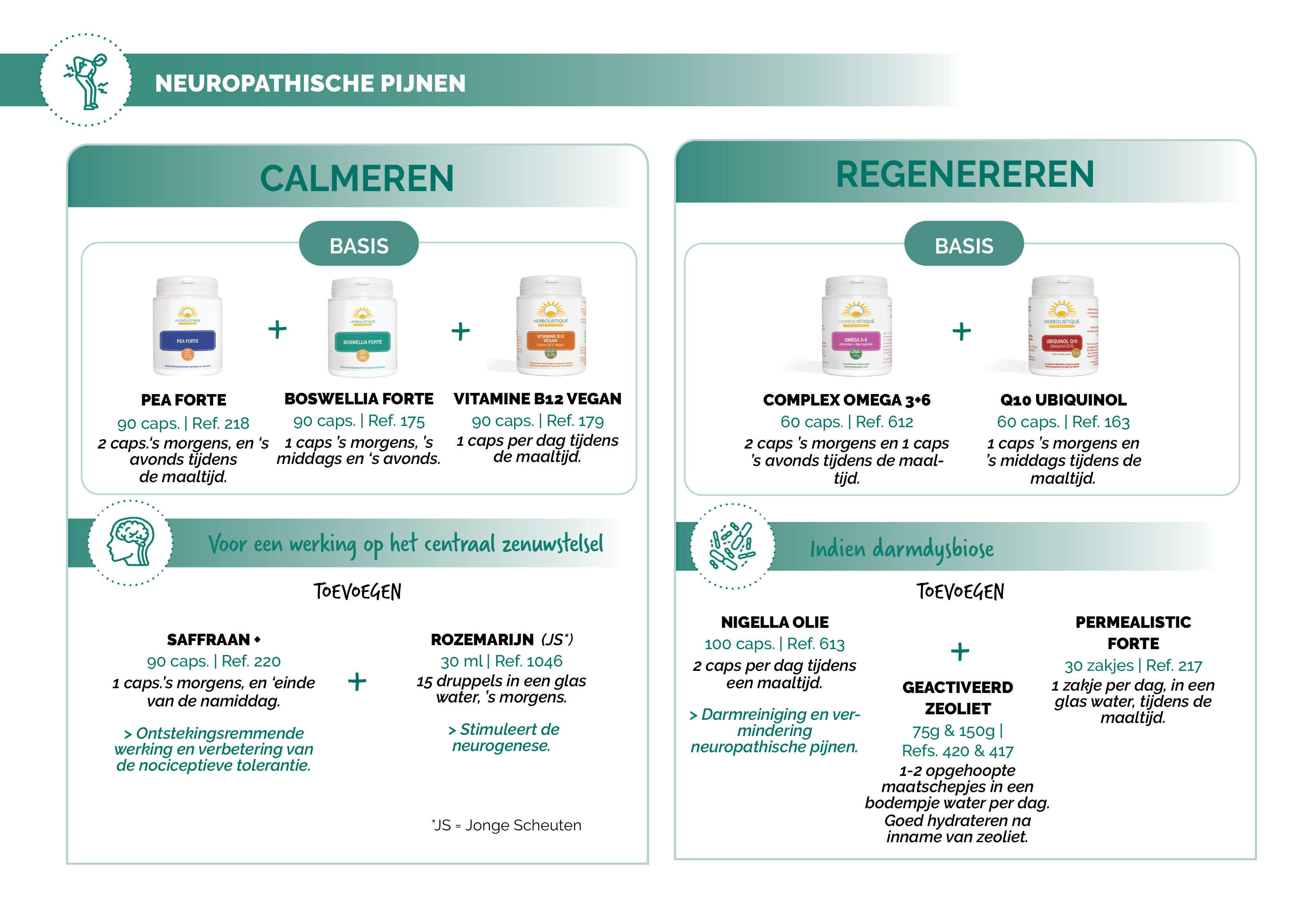
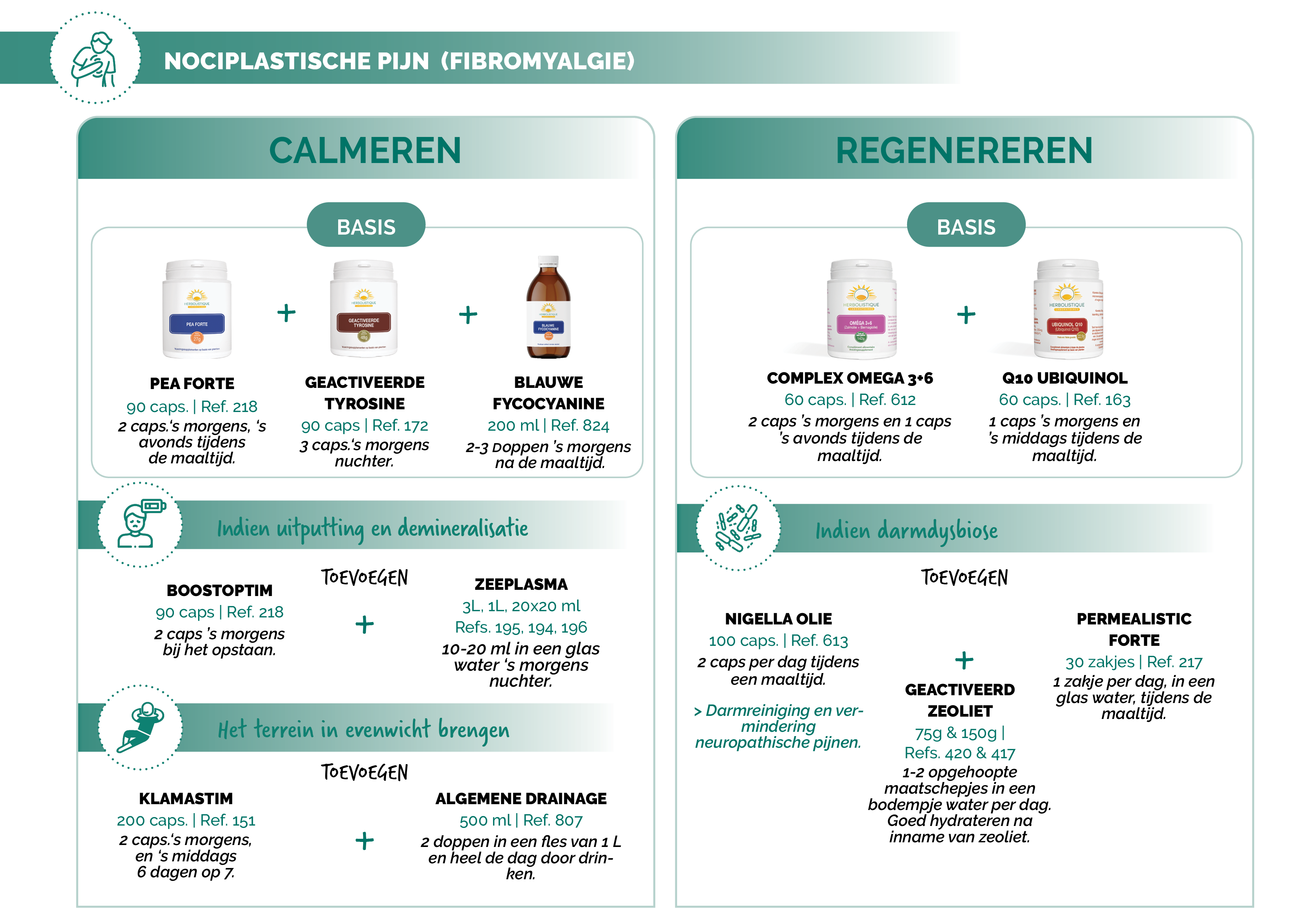
Bronnen:
(1) Masaru Tanaka et al. (2021). Co-Players in Chronic Pain: Neuro-inflammation and the Tryptophan-Kynurenine Metabolic Pathway. Biomedicines 2021, 9, 897.
(2) Fasano, A. (2012). Zonulin and its regulation of intestinal barrier function: The biological door to inflammation, autoimmunity, and cancer. Physiological Reviews, 91(1), 151–175.
(3) Gao, J., Xu, K., Liu, H., Liu, G., Bai, M., Peng, C., … & Li, T. (2018). Impact of the gut microbiota on intestinal immunity mediated by tryptophan metabolism. Frontiers in Cellular and Infection Microbiology, 8, 13.
(4) Calabrese, V., et al. (2021). Neuro-inflammation and neuropathic pain: from mechanisms to therapeutic targets. Journal of Neuro-inflammation, 18, 100.
(5) Ji, R.-R., Nackley, A., Huh, Y., Terrando, N., & Maixner, W. (2016). Neuro-inflammation and central sensitization in chronic and widespread pain. Anesthesiology, 126(3), 830–842.
(6) Banafshe, H. R., Ashtiani, H. R., & Mahmoudi, M. (2018). The role of liver detoxification in pain management. Journal of Pain Research, 11, 1397–1408.
(7) Zhang, Q., et al. (2019). Glutathione metabolism and its implications for pain and neuro-inflammation. Journal of Neurochemistry, 148(2), 167–181.
(8) McEwen, B. S. (2017). Neurobiological and systemic effects of chronic stress. Chronic Stress, 1, 2470547017692328.
(9) Liu, Y., Wang, L., Pan, X., Wang, C., & Wang, Z. (2020). Chronic stress impairs prefrontal cortex synaptic plasticity and cognitive function via NMDA receptor-dependent mechanisms. Neuroscience, 435, 135–145.
(10) Calabrese, F., Rossetti, A. C., Racagni, G., Gass, P., & Riva, M. A. (2021). Stress-induced neuro-inflammation: Mechanisms and new pharmacological targets. Neuroscience & Biobehavioral Reviews, 125, 437–455.
(11) Ji, R. R., Xu, Z. Z., & Gao, Y. J. (2016). Emerging targets in neuro-inflammation-driven chronic pain. Nature Reviews Drug Discovery, 13(7), 533–548.
(12) Schwartz, M., & Quintana, F. J. (2020). The kynurenine pathway in neurological disorders. Trends in Neurosciences, 43(12), 969–978.
(13) Schwartz, M., & Quintana, F. J. (2020). The role of the kynurenine pathway of tryptophan metabolism in inflammation: Mechanisms and therapeutic implications. Journal of Clinical Investigation, 130(5), 1985–1995.
(14) Sheng, J., Liu, S., Wang, Y., Cui, R., & Zhang, X. (2017). The link between depression and chronic pain: Neural mechanisms in the brain. Neural Plasticity, 2017, 9724371.
(15) Bäckryd, E., Tanum, L., Lind, A. L., Larsson, A., & Gordh, T. (2017). Evidence of both systemic inflammation and neuro-inflammation in fibromyalgia patients, as assessed by a multiplex protein panel applied to cerebrospinal fluid and plasma. Journal of Pain Research, 10, 515–525.
(16) Uçeyler, N., Häuser, W., & Sommer, C. (2011). Systematic review with meta‐analysis: cytokines in fibromyalgia syndrome. Rheumatology, 50(3), 534–543.
(17) Loggia, M. L., Berna, C., Kim, J., Cahalan, C. M., Gollub, R. L., Wasan, A. D., Harris, R. E., Edwards, R. R., & Napadow, V. (2015). Disrupted brain circuitry for pain-related reward/punishment in fibromyalgia. Brain, Behavior, and Immunity, 52, 149–158.
(18) Schäfers, M., Svensson, C. I., Sommer, C., & Sorkin, L. S. (2003). Tumor necrosis factor-alpha induces mechanical allodynia after spinal nerve ligation by activation of p38 MAPK in primary sensory neurons. Pain, 105(1–2), 205–213.
(19) Haroon, E., Fleischer, C. C., Felger, J. C., Chen, X., Woolwine, B. J., & Miller, A. H. (2016). Conceptual convergence: increased inflammation is associated with increased basal ganglia glutamate in patients with major depression. Molecular Psychiatry, 21(10), 1351–1357.
(20) Ridker, P. M., Hennekens, C. H., Buring, J. E., & Rifai, N. (2000). C-reactive protein and other markers of inflammation in the prediction of cardiovascular disease in women. The New England Journal of Medicine, 342(12), 836–843.
(21) Tóthová, C., Nagy, O., & Kovac, G. (2022). Serum protein electrophoresis and its diagnostic use in clinical practice: A review. Frontiers in Veterinary Science, 9, 853482.
(22) Calder, P. C. (2011). Fatty acids and inflammation: The cutting edge between food and pharma. Volume 668, Supplement 1, Pages S50-S58
(23) Ganz, T., & Nemeth, E. (2021). Iron homeostasis in host defence and inflammation. Nature Reviews Immunology, 21(1), 26–36.
(24) Ho, P. I., Ortiz, D., Rogers, E., & Shea, T. B. (2002). Multiple aspects of homocysteine neurotoxicity: glutamate excitotoxicity, kinase hyperactivation and DNA damage. Journal of Neuroscience Research, 70(5), 694–702.
(25) Fasano, A. (2020). Zonulin and its regulation of intestinal barrier function: the biological door to inflammation, autoimmunity, and cancer. Cellular and Molecular Gastroenterology and Hepatology, 10(1), 39–46.
(26) Ghosh, S. S., Wang, J., Yannie, P. J., & Ghosh, S. (2020). Intestinal barrier dysfunction, LPS translocation, and disease development. Journal of Endocrinology, 247(1), R13–R34.
(27) Schertzer, J. D., & Klip, A. (2021). Host immune activation by LPS through TLR4–A bridge to insulin resistance and metabolic disease. Cell Metabolism, 33(4), 659–660.
(28) Lang Illievich, K., Klivinyi, C., Lasser, C., Brenna, C. T. A., Szilagyi, I. S., & Bornemann Cimenti, H. (2023). Palmitoylethanolamide in the treatment of chronic pain: A systematic review and meta-analysis of double-blind randomized controlled trials. Nutrients, 15(6), 1350.
(29) Klipinyi, C., Lasser, C., & Bornemann Cimenti, H.-C. (2022). Central and peripheral effects of ultramicronized palmitoylethanolamide in healthy subjects. Nutrients, 14(19), 4084.
(30) Grzanna, R., Lindmark, L., & Frondoza, C. G. (2005). Ginger—An herbal medicinal product with broad anti-inflammatory actions. Journal of Medicinal Food, 8(2), 125–132.
(31) Tripathi, S., Bruch, D., & Kittur, D. S. (2008). Ginger extract inhibits LPS induced macrophage activation and function. BMC Complementary and Alternative Medicine, 8, 1.
(32) Infantino, M., & Di Giacomo, R. (2020). Clinical trials on pain-lowering effect of ginger: A narrative review. Acta Bio Medica: Atenei Parmensis, 91(11-S), e2020013.
(33) Dedov, V. N., Tran, V. H., Duke, C. C., & Ginger, S. (2002). Gingerols: A novel class of vanilloid receptor (VR1) agonists. British Journal of Pharmacology, 137(6), 793–798.
(34) Rahmani, A. H., Shabrmi, F. M. A., & Aly, S. M. (2014). Active ingredients of ginger as potential candidates in the prevention and treatment of diseases via modulation of biological activities. International Journal of Physiology, Pathophysiology and Pharmacology, 6(2), 125.
(35) Bridges, A. R., McCracken, L. M., & Droney, J. (2022). Omega-3 fatty acids and chronic pain: A review of the current evidence. Journal of Pain Research, 15, 1581–1594.
(36) Romay, C., Armesto, J., Remirez, D., González, R., Ledón, N., & García, I. (2003). Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflammopharmacology, 11(3), 255–265.
(37) Benedetti, S., Benvenuti, F., Scoglio, S., & Canestrari, F. (2020). Clinical evaluation of the anti-inflammatory and pain-relieving effects of a natural extract of spirulina: A pilot study on osteoarthritis. Clinical Nutrition ESPEN, 35, 159–165.
(38) Lopresti, A. L., & Drummond, P. D. (2014). Saffron (Crocus sativus) for depression: A systematic review of clinical studies and examination of underlying antidepressant mechanisms of action. Human Psychopharmacology: Clinical and Experimental, 29(6), 517–527.
(39) Shafiee, M., Arekhi, S., Omranzadeh, A., & Sahebkar, A. (2018). Saffron in the treatment of depression, anxiety and other mental disorders: Current evidence and potential mechanisms of action. Journal of Affective Disorders, 227, 330–337.
Document uitsluitend bestemd voor de gezondheidsprofessional.